
Effects of Exogenous Organic Acids on the Soil Metabolites and Microbial Communities of Panax notoginseng from the Forest Understory

Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Experimental Design and Sample Collection
2.3. Fresh Weight and Edaphic Factors
2.4. DNA Extraction, PCR Amplification, and Sequencing
2.5. Metabolite Profiling from Rhizosphere Samples
2.6. Statistical Analysis
3. Results
3.1. Analysis of the Fresh Weight of Sanqi and Soil Physicochemical Properties under Organic Acid Conditions
3.2. α- and β-Diversity of Bacteria and Fungi Analysis
3.3. Dominant Phyla and Genus of Bacteria and Fungi
3.4. Complexity and Stability of Bacterial and Fungal Network
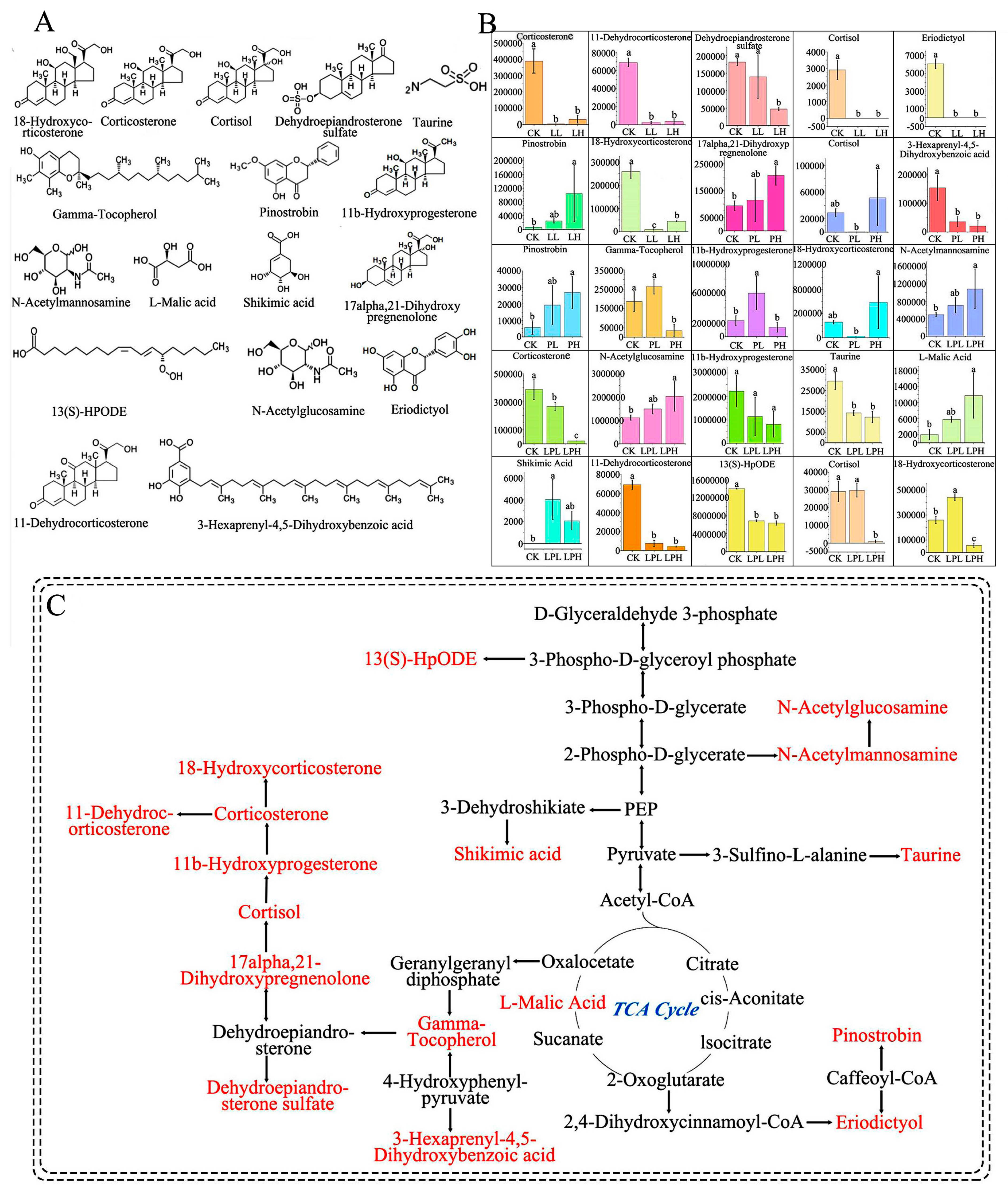
3.5. Differential Metabolites (DMs) in Sanqi Soils
3.6. Correlation of Soil Microbes with DMs
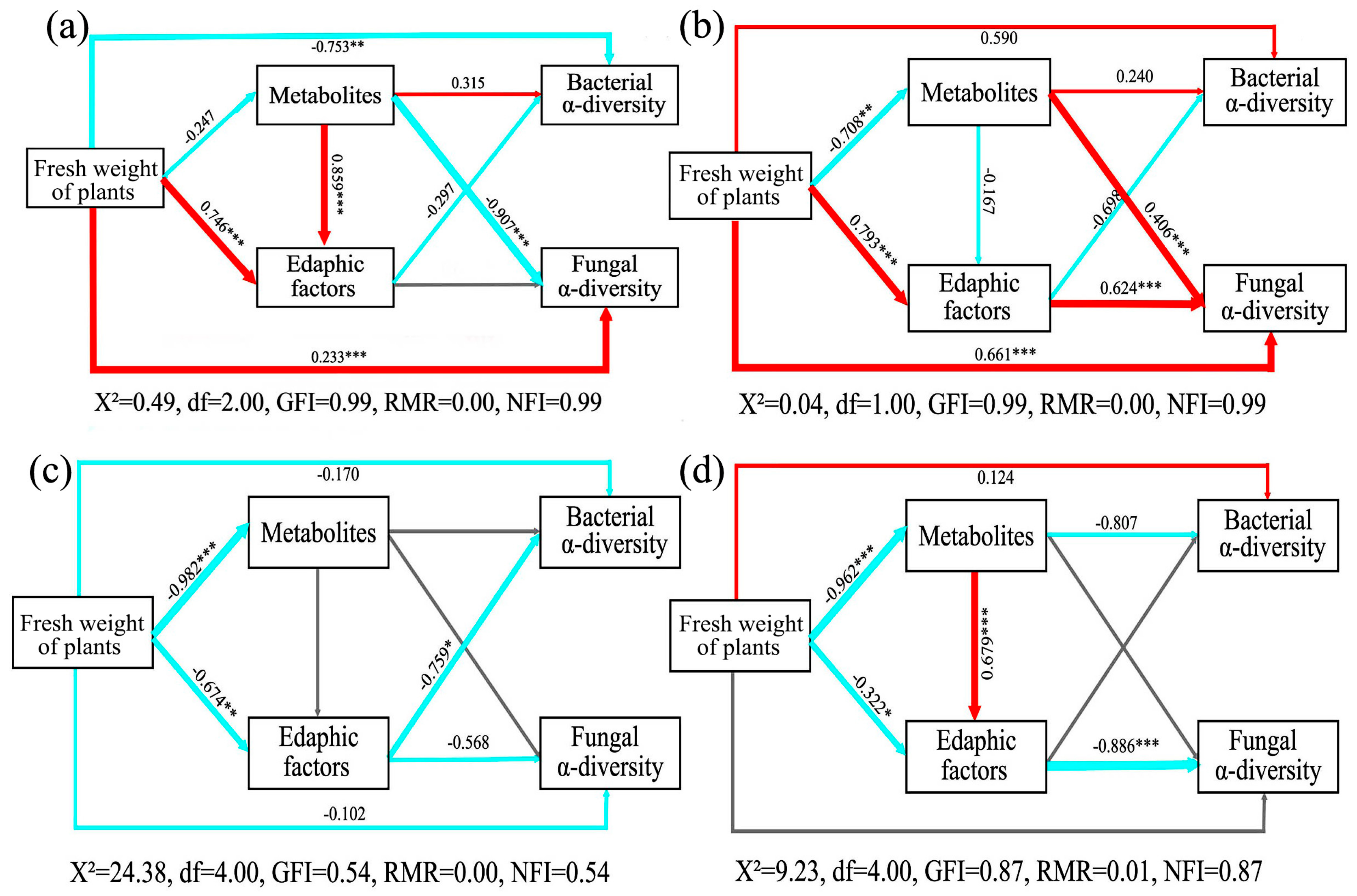
3.7. Structural Equation Model (SEM) Analysis
4. Discussion
4.1. Organic Acid Concentrations of >250 mg/kg Reduced Sanqi Growth and Soil’s Physicochemical Properties
4.2. Organic Acid Concentrations of >250 mg/kg Increased Microbial α-Diversity
4.3. Different Organic Acids Affect the Dominant Flora in Sanqi Soils Differently
4.4. Concentration of Organic Acids of >250 mg/kg Reduced the Complexity and Stability of Microbial Network
4.5. Different Types of Organic Acids Changed Soil DMs and Metabolic Pathways
4.6. Organic Acid Changed the Relationship among SANQI, Edaphic Factors, Metabolites, and Soil Microbes
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Zhang, Z.Y.; Lin, W.X. Continuous cropping obstacle and allelopathic autotoxicity of medicinal plants. Chin. J. Eco-Agric. 2009, 17, 189–196. [Google Scholar] [CrossRef]
- Li, S.L.; Jing, Y.B.; Mao, J.H.; Li, Y.P.; Li, R.B. Effects of exogenous phenolic acids on growth and photosynthesis characteristics of Santalum album seedlings. J. West China For. Sci. 2020, 49, 104–108. [Google Scholar] [CrossRef]
- Li, R.; Shen, Z.Z.; Sun, L.; Zhang, R.F.; Fu, L.; Deng, X.H.; Shen, Q.R. Novel soil fumigation method for suppressing cucumber Fusarium wilt disease associated with soil microflora alterations. Appl. Soil Ecol. 2016, 101, 28–36. [Google Scholar] [CrossRef]
- Xiang, W.; Chen, J.; Zhang, F.; Huang, R.; Li, L. Autotoxicity in Panax notoginseng of root exudates and their allelochemicals. Front. Plant Sci. 2022, 13, 1020626. [Google Scholar] [CrossRef]
- Gong, J.T.; Cheng, X.Y.; Sun, M.; Wu, L.J.; Zhang, Z.L. Effects of three kinds of saponins on seed germination and seedling growth of Panax notoginseng. Acta Agric. Univ. Jiangxiensis 2015, 37, 988–993. [Google Scholar] [CrossRef]
- You, P.J.; Zhang, Y.; Wang, W.Q.; Zhang, Z.L.; Yang, J.Z.; Yin, L.M. Allelopathic effects of continuous cropping soil of Panax notoginseng on seed and seedling of some vegetables. Mod. Chin. Med. 2009, 5, 12–13. [Google Scholar]
- An, N.B. Study on the Chemotaxis of Main Ginseng Disease to Saccharides and Amino Acids of Ginseng Root Exudates. Master’s Thesis, Jilin Agricultural University, Changchun, China, 2017. [Google Scholar]
- Wu, L.J.; Liu, J.; Wang, W.Y.; Dai, D.; Cheng, X.Y.; Zhang, Z.L.; Yang, Y.J. Identification and content determination of phenolic acids of rhizosphere soil of Panax notoginseng. World Sci. Technol.-Mod. Tradit. Chin. Med. 2014, 16, 825–829. [Google Scholar] [CrossRef]
- Hei, J.Y.; Wang, S.; He, X.H. Effects of exogenous organic acids on the growth, edaphic factors, soil extracellular enzymes, and microbiomes predict continuous cropping obstacles of Panax notoginseng from the forest understorey. Plant Soil 2023, 1–18. [Google Scholar] [CrossRef]
- Zheng, Y.B.; Li, M. Autotoxicity of phenolic acids in root exudates of Andrographis paniculata (Burm. F.) Nees. Allelopath. J. 2018, 45, 153–162. [Google Scholar] [CrossRef]
- Chen, L.; Zhang, M.L.; Xin, M.Y.; Li, J.D. Effects of exogenous phenolic acids on allelopathy of potted soybean seedlings. J. Agric. Sci. Technol. 2015, 16, 1151. [Google Scholar] [CrossRef]
- Li, L.; Jiang, J.L. Research advances in allelopathic autotoxicity and continuous cropping obstacle of American ginseng. Mol. Plant Breed. 2018, 16, 4436–4443. [Google Scholar] [CrossRef]
- Chen, L.C.; Liao, L.P.; Wang, S.L.; Huang, Z.Q. Effect of exotic toxin on the nutrition of woodland soil. Chin. J. Ecol. 2002, 21, 19–22. [Google Scholar]
- Shen, Q.R.; Yu, T.Y.; Zhu, H.T. The effect of added phenolic acids on soil enzyme activities and nutrients. Plant Nutr. Fertil. Sci. 2006, 12, 845–849. [Google Scholar] [CrossRef]
- Wang, Y.P.; Wang, H.T.; Xu, T.; Ni, G.P.; Jiang, Y.Z. Effects of exogenous phenolic acid on soil nutrient availability and enzyme activities in a poplar plantation. Ying Yong Sheng Tai Xue Bao 2013, 24, 667–674. [Google Scholar] [CrossRef]
- Wang, B. Study on the Mechanism of Exogenous Phenolic Acids on Bata Grape Plan Growth and Rhizosphere SOIL microbes. Doctoral Thesis, Shenyang Agricultural University, Shenyang, China, 2016. [Google Scholar]
- Tan, X.M. Effects of Alternative Continuous Cropping of Poplar Cultivars Alternative on Soil Ecological Environment and Allelopathy of Exogenous Phenolic Acid. Master’s Thesis, Shandong Agricultural University, Taian, China, 2008. [Google Scholar]
- Shen, H.; Yan, W.H.; Yang, X.Y.; He, X.H.; Wang, X.; Zhang, Y.T.; Wang, B.; Xia, Q.Y. Co-occurrence network analyses of rhizosphere soil microbial PLFAs and metabolites over continuous cropping seasons in tobacco. Plant Soil 2020, 452, 119–135. [Google Scholar] [CrossRef]
- Yang, Y.; Li, H.L.; Ma, K.L.; Yu, F.F.; Niu, S.Q. Effect of continuous cropping on the physicochemical properties, microbial activity, and community characteristics of the rhizosphere soil of codonopsis pilosula. Environ. Sci. 2023, 44, 6387–6398. [Google Scholar] [CrossRef]
- Were, E.; Schöne, J.; Viljoen, A.; Rasche, F. Phenolics mediate suppression of Fusarium oxysporum f. sp. cubense TR4 by legume root exudates. Rhizosphere 2022, 21, 100459. [Google Scholar] [CrossRef]
- Einhellig, F.A. Interactions involving allelopathy in cropping systems. Agron. J. 1996, 88, 886–893. [Google Scholar] [CrossRef]
- Chen, P.; Wang, Y.Z.; Liu, Q.Z.; Zhang, Y.T.; Li, X.Y.; Li, H.Q.; Li, W.H. Phase changes of continuous cropping obstacles in strawberry (Fragaria×ananassa Duch.) production. Appl. Soil Ecol. 2020, 155, 103626. [Google Scholar] [CrossRef]
- Shi, R.; Wang, S.; Xiong, B.J.; Gu, H.Y.; Wang, H.L.; Ji, C.; Jia, W.J.; Abraham, R.H.; Zhen, W.J.; Jiftah, B.A.; et al. Application of bioorganic fertilizer on Panax notoginseng improves plant growth by altering the rhizosphere microbiome structure and metabolism. Microorganisms 2022, 10, 275. [Google Scholar] [CrossRef] [PubMed]
- Yu, P.; He, X.; Baer, M.; Beirinckx, S.; Tian, T.; Moya, Y.A.; Hochholdinger, F. Plant favones enrich rhizosphere Oxalobacteraceae to improve maize performance under nitrogen deprivation. Nat. Plants 2021, 7, 481–499. [Google Scholar] [CrossRef]
- Jia, W.J.; Hei, J.Y.; He, X.H.; Wang, S. Altitude rather than season and slope aspect has the greatest effect on the bacterial communities in subtropical forests in Yunnan, China. Plant Soil 2023, 1–18. [Google Scholar] [CrossRef]
- Jiao, S.; Chen, W.M.; Wang, J.L.; Du, N.N.; Li, Q.P.; Wei, G.H. Soil microbiomes with distinct assemblies through vertical soil profiles drive the cycling of multiple nutrients in reforested ecosystems. Microbiome 2018, 6, 146. [Google Scholar] [CrossRef]
- Liao, H.; Hao, X.L.; Qin, F.; Delgado-Baquerizo, M.; Liu, Y.R.; Zhou, J.Z.; Cai, P.; Chen, W.L.; Huang, Q.Y. Microbial autotrophy explains large-scale soil CO2 fixation. Glob. Chang. Biol. 2023, 29, 231–242. [Google Scholar] [CrossRef]
- Xun, W.B.; Liu, Y.P.; Li, W.; Ren, Y.; Xiong, W.; Xu, Z.H.; Zhang, N.; Miao, Y.Z.; Shen, Q.R.; Zhang, R.F. Specialized metabolic functions of keystone taxa sustain soil microbiome stability. Microbiome 2021, 9, 35. [Google Scholar] [CrossRef]
- Balke, N.E. Effects of allelochemicals on mineral uptake and associated physiological processes. ACS Symp. Ser. 1985, 268, 161–178. [Google Scholar] [CrossRef]
- Zhang, J.H.; Mao, Z.Q.; Wang, L.Q.; Wang, L.Q.; Shu, H.R. Effect of phloridzin on physiological characteristics of Malus hupehensis Rehd. Seedlings. Sci. Agric. Sin. 2007, 40, 492–498. [Google Scholar]
- Yuan, N.; Zheng, R.H.; Gu, X.P.; Wu, X.L.; Yue, J.J. Allelopathic effect of phenolic acids on Phyllostachys edulis seedlings. Nanjing For. Univ. 2010, 34, 29–33. [Google Scholar] [CrossRef]
- Lehman, M.E.; Blum, U.; Gerig, T.M. Simultaneous effects of ferulic and p-coumaricacids on cucumber leaf expansion in split-root experiments. J. Chem. Ecol. 1994, 20, 1773–1782. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.J.; Li, T.Y.; Ye, C.; Lu, R.C.; Liu, Y.X.; Huang, H.C.; He, X.H.; Yan, M.; Zhu, S.S. Leaching alleviates phenol–mediated root rot in Panax notoginseng by modifying the soil microbiota. Plant Soil 2021, 468, 491–507. [Google Scholar] [CrossRef]
- Bardhan, S.; Jose, S.; Jenkins, M.A.; Webster, C.R.; Udawatta, R.P.; Stehn, S.E. Microbial community diversity and composition across a gradient of soil acidity in spruce-fir forests of the southern Appalachian Mountains. Appl. Soil Ecol. 2012, 61, 60–68. [Google Scholar] [CrossRef]
- Wei, H.J.; Ding, J.; Zhang, J.M.; Yang, W.; Wang, Y.Q.; Liu, T.Z. Changes in soil fungal community structure under bermudagrass turf in response to traffic stress. Acta Pratacul. Sin. 2022, 31, 102–112. [Google Scholar] [CrossRef]
- Yelle, D.J.; Ralph, J.; Ralph, J.; Lu, F.; Hammel, K.E. Evidence for cleavage of lignin by a brown rot Basidiomycete. Environ. Microbiol. 2008, 10, 1844–1849. [Google Scholar] [CrossRef] [PubMed]
- Miao, C.P.; Mi, Q.L.; Qiao, X.G.; Zheng, Y.K.; Chen, Y.W.; Xu, L.H.; Guan, H.L.; Zhao, L.X. Rhizospheric fungi of Panax notoginseng: Diversity and antagonism to host phytopathogens. J. Ginseng Res. 2016, 40, 127–134. [Google Scholar] [CrossRef] [PubMed]
- Shen, J.; Yang, G.T.; Liu, D.J. Influence of wood vinegar and pine needle on soil improvement of blueberry. Soil 2014, 46, 325–329. [Google Scholar] [CrossRef]
- Han, Q.; Ma, Q.; Chen, Y.; Tian, B.; Xu, L.X.; Bai, Y.; Chen, W.F.; Li, X. Variation in rhizosphere microbial communities and its association with the symbiotic efficiency of rhizobia in soybean. ISME J. 2020, 14, 1915–1928. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.M.; Cheng, Y.X.; Ma, Y.N.; Chen, C.J.; Xu, F.R.; Dong, X. Role of phenolic acids from the rhizosphere soils of Panax notoginseng as a double-edge sword in the occurrence of root–rot disease. Molecules 2018, 23, 819. [Google Scholar] [CrossRef]
- Perrone, G.; Susca, A. Penicillium species and their associated Mycotoxins. Methods Mol. Biol. 2017, 1542, 107–119. [Google Scholar] [CrossRef]
- Xu, A.Y.; Hu, Z.H.; Jiao, S.; Xu, Q.; Ma, L.L.; Chen, J. Depth-dependent effects of tree species identity on soil microbial community characteristics and multifunctionality. Sci. Total Environ. 2023, 878, 162972. [Google Scholar] [CrossRef]
- Shi, S.J.; Nuccio, E.E.; Shi, Z.J.; He, Z.L.; Zhou, J.Z.; Firestone, M.K. The interconnected rhizosphere: High network complexity dominates rhizosphere assemblages. Ecol. Lett. 2016, 19, 926–936. [Google Scholar] [CrossRef]
- Yuan, M.M.; Guo, X.; Wu, L.W.; Zhang, Y.; Xiao, N.J.; Ning, D.L.; Zhou, S.; Zhou, X.S.; Wu, L.Y.; Yang, Y.F.; et al. Climate warming enhances microbial network complexity and stability. Nat. Clim. Chang. 2021, 11, 343–348. [Google Scholar] [CrossRef]
- Hu, X.J.; Liu, J.J.; Wang, X.Z.; Yu, Z.H.; Yao, Q.; Jin, J.; Liu, X.B.; Wang, G.H. Dramatic changes in bacterial co-occurrence patterns and keystone taxa responses to cropping systems in Mollisols of Northeast China. Arch. Agron. Soil Sci. 2021, 67, 426–434. [Google Scholar] [CrossRef]
- Liu, C.H.; He, H.; He, X.G.; Liu, K.; Shao, X.H.; Lai, D.; Kuang, S.Z.; Xiao, W.Q. Analysis of differential metabolites and bacterial community structure in soils of a pineapple orchard in different continuous-cropping years. Biotechnol. Bull. 2021, 37, 162–175. [Google Scholar] [CrossRef]
- Wang, D.Y. Study on Environmental Behavior of Natural Steroid Hormones in Soils and Its Risk Control. Doctoral Dissertation, China University of Mining and Technology, Beijing, China, 2015. [Google Scholar]
- Mandal, S.M.; Chakraborty, D.; Dey, S. Phenolic acids act as signaling molecules in plant-microbe symbioses. Plant Signal. Behav. 2010, 5, 359–368. [Google Scholar] [CrossRef]
- Zeeshan Ul Haq, M.; Yu, J.; Yao, G.; Yang, H.; Iqbal, H.A.; Tahir, H.; Wu, Y. A systematic review on the continuous cropping obstacles and control strategies in medicinal plants. Int. J. Mol. Sci. 2023, 24, 12470. [Google Scholar] [CrossRef]
- Blum, U.; Shafer, S.R. Microbial-populations and phenolic acids in soil. Soil Biol. Biochem. 1988, 20, 793–800. [Google Scholar] [CrossRef]
- Garbeva, P.; Van Veen, J.A.; van Elsas, J.D. Microbial diversity in soil: Selection of microbial populations by plant and soil type and implications for disease suprressiveness. Annu. Rev. Phytopathol. 2004, 42, 243–270. [Google Scholar] [CrossRef] [PubMed]
- Lareen, A.; Burton, F.; Schafer, P. Plant root-microbe communication in shaping root microbiomes. Plant Mol. Biol. 2016, 90, 575–587. [Google Scholar] [CrossRef] [PubMed]
- Lynch, J.M.; Whipps, J.M. Substrate flow in the rhizosphere. Plant Soil 1990, 129, 1–10. [Google Scholar] [CrossRef]
- Knief, C.; Delmotte, N.; Vorholt, J.A. Bacterial adaptation to life in association with plants-A proteomic perspective from culture to in situ conditions. Proteomics 2011, 11, 3086–3105. [Google Scholar] [CrossRef] [PubMed]
- Chaparro, J.M.; Badri, D.V.; Vivanco, J.M. Rhizosphere microbiome assemblage is affected by plant development. ISME J. 2014, 8, 790–803. [Google Scholar] [CrossRef] [PubMed]
- Sasse, J.; Martinoia, E.; Northen, T. Feed your friends: Do plant exudates shape theroot microbiome? Trends Plant Sci. 2018, 23, 25–41. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| AVD of Bacterial Community (15 Days) | AVD of Bacterial Community (30 Days) | AVD of Fungal Community (15 Days) | AVD of Fungal Community (30 Days) | |
|---|---|---|---|---|
| CK | 0.66 ± 0.09 bc | 0.651 ± 0.01 b | 0.88 ± 0.05 a | 0.73 ± 0.03 ab |
| L (<250 mg/kg) | 0.78 ± 0.14 ab | 0.666 ± 0.19 b | 0.65 ± 0.02 b | 0.60 ± 0.09 c |
| L (>250 mg/kg) | 0.62 ± 0.06 c | 0.818 ± 0.12 abc | 0.60 ± 0.03 b | 0.80 ± 0.03 a |
| LP (<250 mg/kg) | 0.79 ± 0.04 ab | 0.693 ± 0.06 bc | 0.68 ± 0.05 b | 0.69 ± 0.05 b |
| LP (>250 mg/kg) | 0.82 ± 0.15 a | 0.875 ± 0.0 a | 0.66 ± 0.06 b | 0.80 ± 0.02 a |
| P (<250 mg/kg) | 0.76 ± 0.04 abc | 0.686 ± 0.0 bc | 0.59 ± 0.05 b | 0.73 ± 0.05 ab |
| P (>250 mg/kg) | 0.73 ± 0.12 abc | 0.848 ± 0.06 ab | 0.64 ± 0.1 b | 0.78 ± 0.01 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hei, J.; Li, Y.; Wang, Q.; Wang, S.; He, X. Effects of Exogenous Organic Acids on the Soil Metabolites and Microbial Communities of Panax notoginseng from the Forest Understory. Agronomy 2024, 14, 601. https://doi.org/10.3390/agronomy14030601
Hei J, Li Y, Wang Q, Wang S, He X. Effects of Exogenous Organic Acids on the Soil Metabolites and Microbial Communities of Panax notoginseng from the Forest Understory. Agronomy. 2024; 14(3):601. https://doi.org/10.3390/agronomy14030601
Chicago/Turabian StyleHei, Jingying, Yue Li, Qiong Wang, Shu Wang, and Xiahong He. 2024. "Effects of Exogenous Organic Acids on the Soil Metabolites and Microbial Communities of Panax notoginseng from the Forest Understory" Agronomy 14, no. 3: 601. https://doi.org/10.3390/agronomy14030601